Slipping into a K-hole - by Andrew R. Gallimore Slipping into a K-hole
Length: • 6 mins
Annotated by Zakhar
Zakhar: This article explains a possible mechanism by which ketamine induces a dissociative state known as a "K-hole". It suggests that ketamine binds to glutamate-gated ion channels known as NMDA receptors in the cortex, which can either lead to increased neural activity at low doses or suppressed neural activity at higher doses. This creates a three-level plateau of effects, including a low-level psychedelic effect, a dissociative effect, and an anaesthetised state.
The Mechanism of Ketamine’s “Dissociated” State
From the earliest clinical studies in humans in the 1960s, subjects injected with ketamine reported feeling detached from their bodies, floating in outer space, and completely disconnected — dissociated — from the outside world. An entirely new class of psychoactive molecule was born — the dissociative anaesthetics — with ketamine as its founding member.
Until now, the mechanism by which ketamine elicits this state, in which the user becomes temporarily disconnected from the environment, whilst experiencing vivid, internally-generated visions, was unclear. However, a new paper published in Nature Neuroscience this week reveals a possible mechanism. To understand how it works, we need to consider how the brain constructs the experienced world and how this world is connected to the environment.
See: https://doi.org/10.1038/s41593-022-01203-5

The experienced world is a model of the environment built using patterns of cortical activity. This model is used to predict patterns of information arriving via the senses. Successfully predicted sensory information is extinguished (filtered out), whereas unpredicted or surprising information generates error signals which pass into the cortex and are used to update the model. The magnitude of an error signal is essentially the difference between the predicted and actual received sensory information. So, the construction and maintenance of the world model is a continuous cycle of model testing, effected by the flow of information into and out of the world-building areas of the cortex. See my previous article for more details on how this works:
Some content could not be imported from the original document. View content ↗
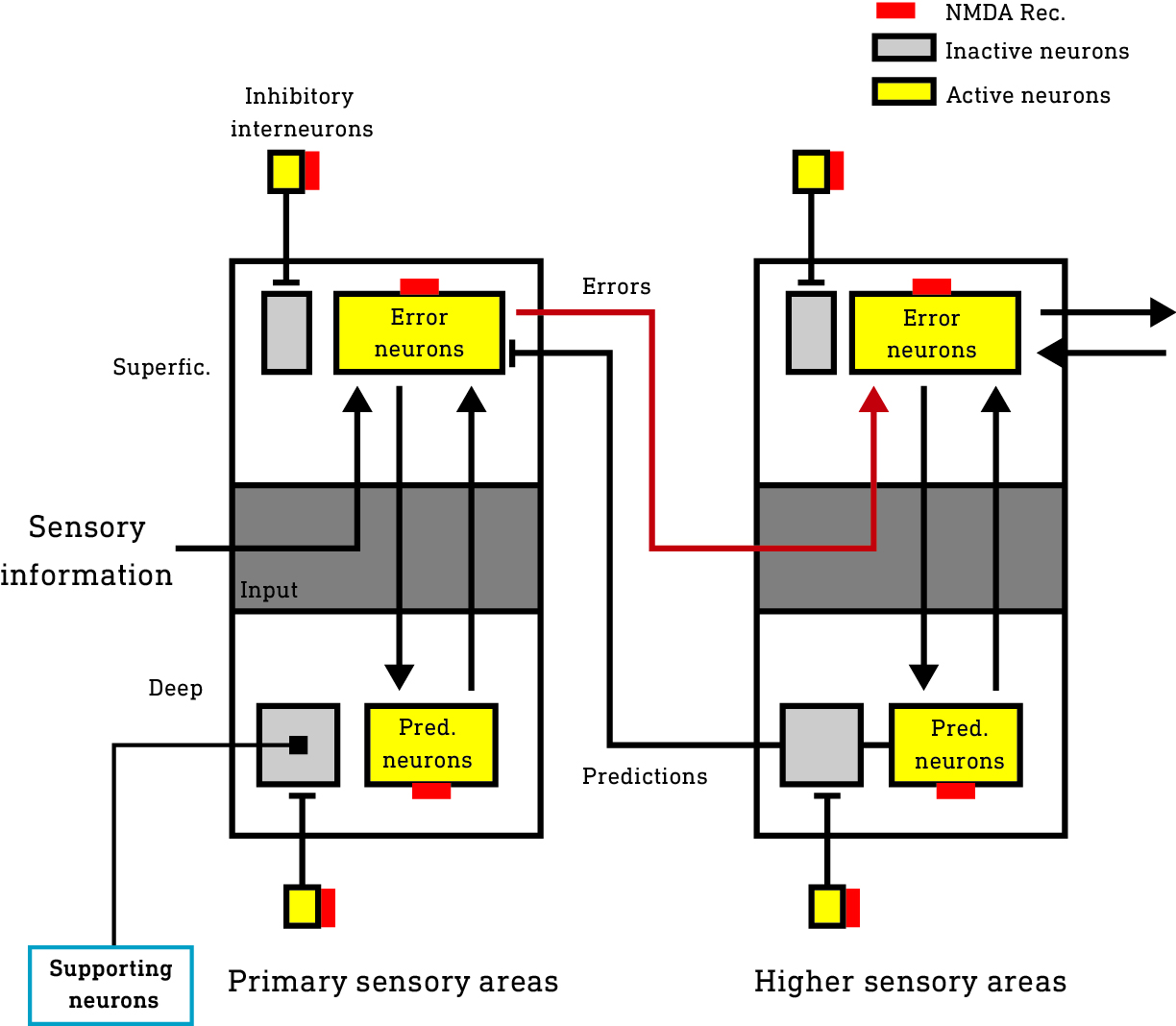
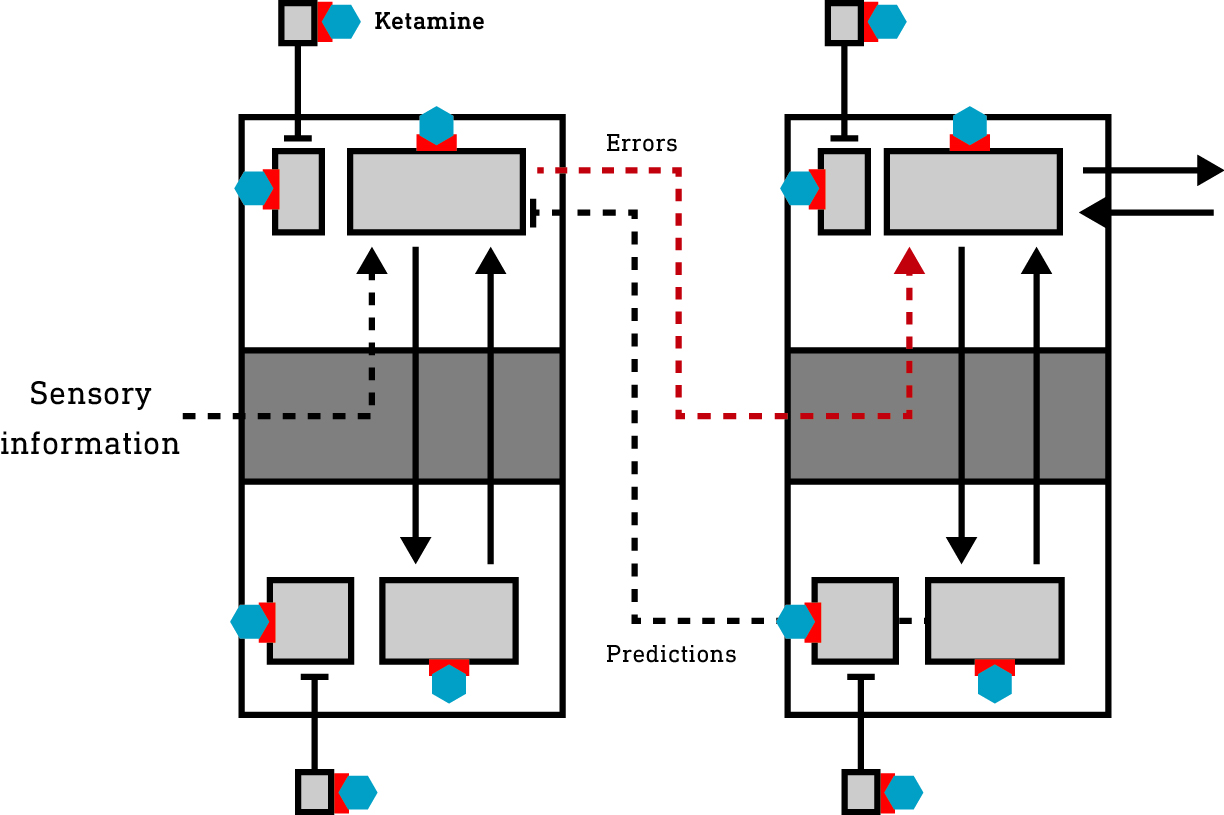
Areas of the cortex form a hierarchy from the lowest — primary — areas that receive information from the sensory apparatus, to higher areas responsible for constructing and maintaining the model you experience as your subjective world. The cortex is also subdivided into six layers, each comprised of a number of types of information-generating neurons and interconnected with the other layers in a tightly regulated pattern. Neurons in the deep layers (layers 5 and 6) are responsible for generating model predictions and transmitting them down the cortical hierarchy to the primary areas. The superficial layer neurons (layers 1-3) are responsible for comparing these predictions to actual sensory information and calculating the prediction errors, which are then sent to neurons in the input layer (layer 4) in the next (higher) level of the hierarchy. So, the prediction errors are broadcast up the cortical hierarchy (with the predictions flowing in the opposite direction) and are used to refine the model until the flow of errors is stemmed (meaning the model is better able to predict the sensory information).

So, it is these neurons responsible for transmitting predictions and error signals that are effectively connected to the environment. Other neurons are important in supporting cortical activity but aren’t directly connected to the environment since they neither generate predictions nor error signals.
Ketamine works by binding to glutamate-gated ion channels known as NMDA receptors in the cortex. Normally, when glutamate binds to this receptor, the channel opens and allows positively-charged ions to flow into the neuron, making it more likely that the neuron will fire. However, crucially, the ion channel is normally blocked by a magnesium ion and will only open fully if glutamate is bound and the magnesium ion has dissociated. Magnesium is more likely to dissociate from the NMDA receptor when the neuron is in a depolarised (“activated”) state.
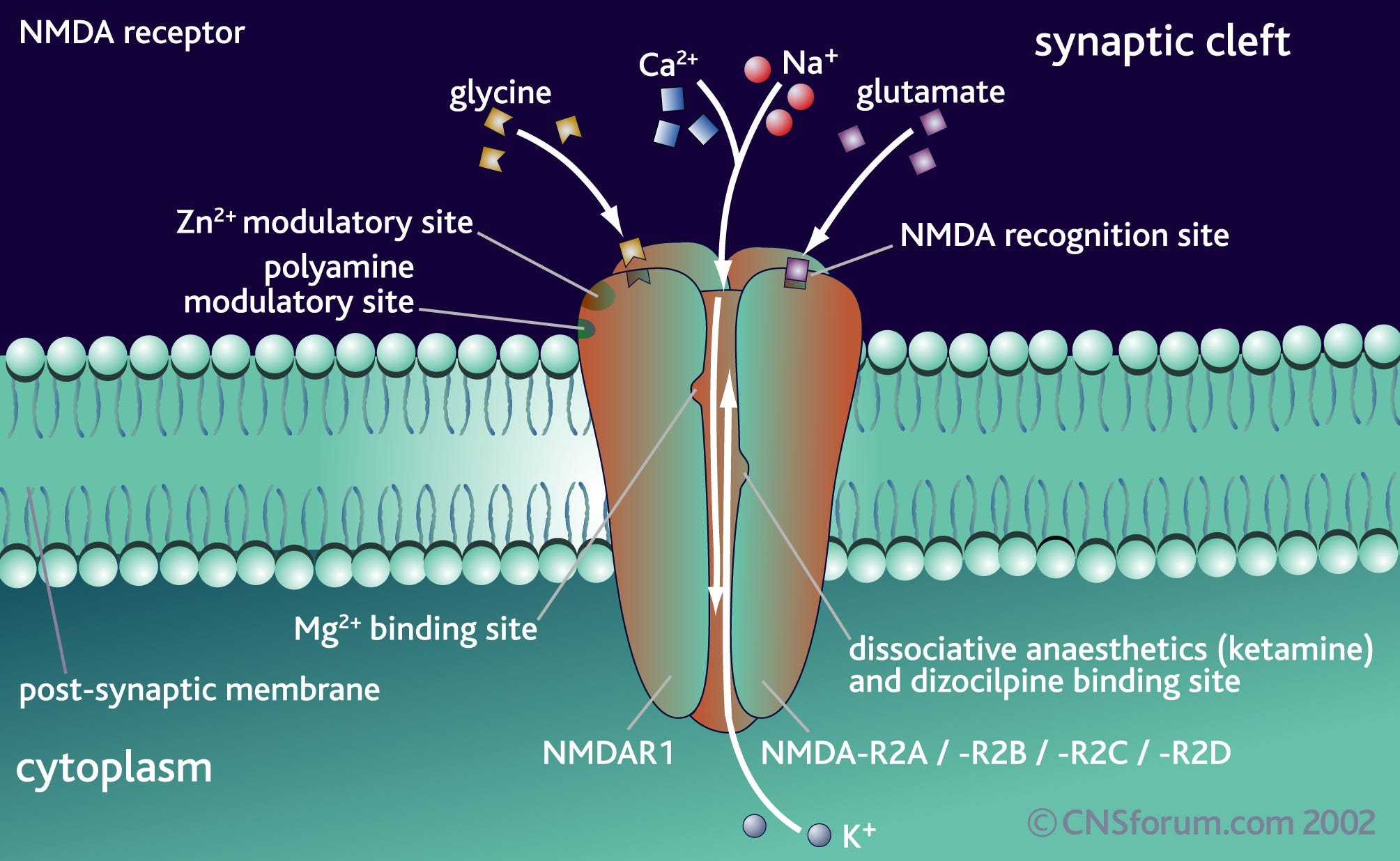
NMDA receptors are found in the membranes of neurons all across the brain, but they have important roles in regulating the activity of, and controlling the flow of information between, neurons that generate the patterns of neural activity that represent your experienced world model. In particular, NMDA receptors are used by neurons that control the flow of information up and down the cortical hierarchy. They are also found on inhibitory interneurons that help to control (by inhibiting) the activity of excitatory neurons across the cortex, as well as on the excitatory neurons themselves.
Ketamine sits in the ion channel part of the NMDA receptor and blocks the flow of ions, even when glutamate is bound. In other words, it acts as an NMDA receptor antagonist. This exhibits what’s known as “use dependence”, which means the ketamine molecule can only bind and block the NMDA ion channel when the magnesium ion has been displaced. This means ketamine has a stronger effect on neurons that are already highly active, since their NMDA receptors will spend more time without the magnesium block when ketamine can bind.
In the basal state (before ketamine reaches the brain), a large population of excitatory neurons (we’ll call them supporting neurons [blue box]) not involved in generating or transmitting predictions/error are suppressed by inhibitory interneurons and largely inactive (in grey).
The inhibitory interneurons tend to fire at a constant high rate and, as such, are highly susceptible to the effects of ketamine. The suppression of these inhibitory neurons releases the previously quiet excitatory supporting neurons in both the deep and superficial layers of the cortex from inhibition — they are disinhibited and become more active. Indeed, at low dose levels, ketamine has been shown to increase neural activity across the cortex. This is experienced as a mild psychedelic effect, not so dissimilar from a low dose of one of the classic 5HT2A agonist psychedelics, which also stimulate cortical activity, albeit by a different mechanism.
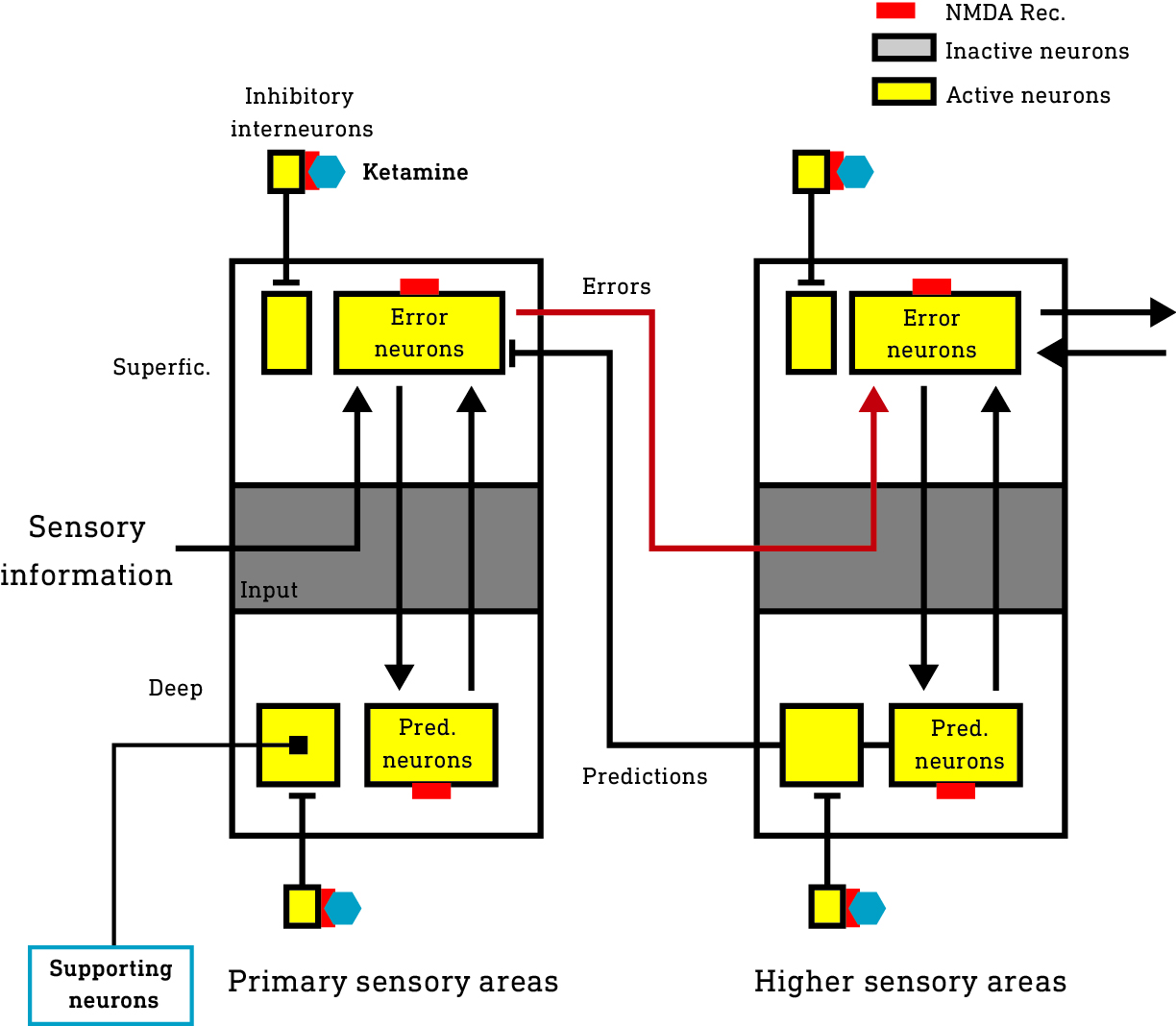
However, as the dosage is increased, a more dissociative effect begins to manifest, in which awareness of the environment is progressively lost, cultivating in a fully disconnected “K-hole” state, which is usually undesirable (but fortunately only likely to persist for 10-20 mins or so). The margin of error seems to be quite narrow and it’s quite easy to slip over the edge into the K-hole, suggestive of a kind of threshold switching behaviour, which is what this new research reveals.
Since the generation of predictions and error signals is a continuous process whenever you’re awake and navigating the environment, these neurons in the deep and superficial layers tend to be highly active and rely partly on NMDA receptors to transmit their signals and maintain their activity. However, unlike the inhibitory interneurons which tend to fire continuously at a high rate, these excitatory neurons are usually active (fire) in intermittent bursts, making them somewhat less susceptible to NMDA blockade. It’s not surprising then that they’re largely unaffected by ketamine until the brain concentration reaches a higher threshold, beyond which they are strongly subdued, which suppresses the generation and propagation of predictions and error signals, effectively disconnecting — dissociating — the cortex from the environment.
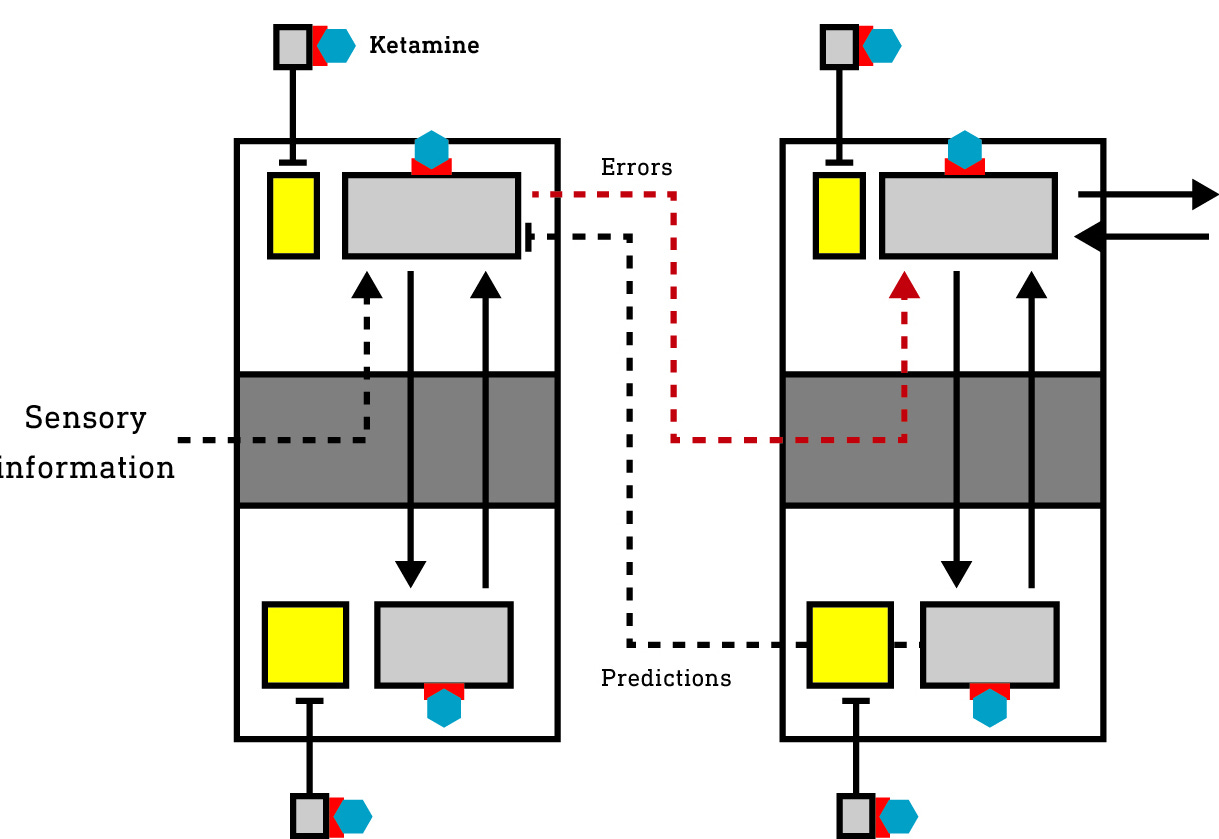
However, the supporting neurons (those not generating predictions or errors) that were activated (disinhibited) by interneuron suppression remain active and constructing a world model, albeit one disconnected from the environment. Since this model isn’t held accountable to sensory testing, novel patterns of activity can emerge, experienced as the dream-like and often confusing K-hole state. It is in this state that highly complex visual worlds unlike the normal waking world, and often populated by entities, can emerge.
As the dosage is increased further, these newly activated supporting neurons also become suppressed by NMDA blockade, cortical activity is more generally suppressed, and a unique anaesthetised state ensues. See my previous article for more details on this state:
Some content could not be imported from the original document. View content ↗
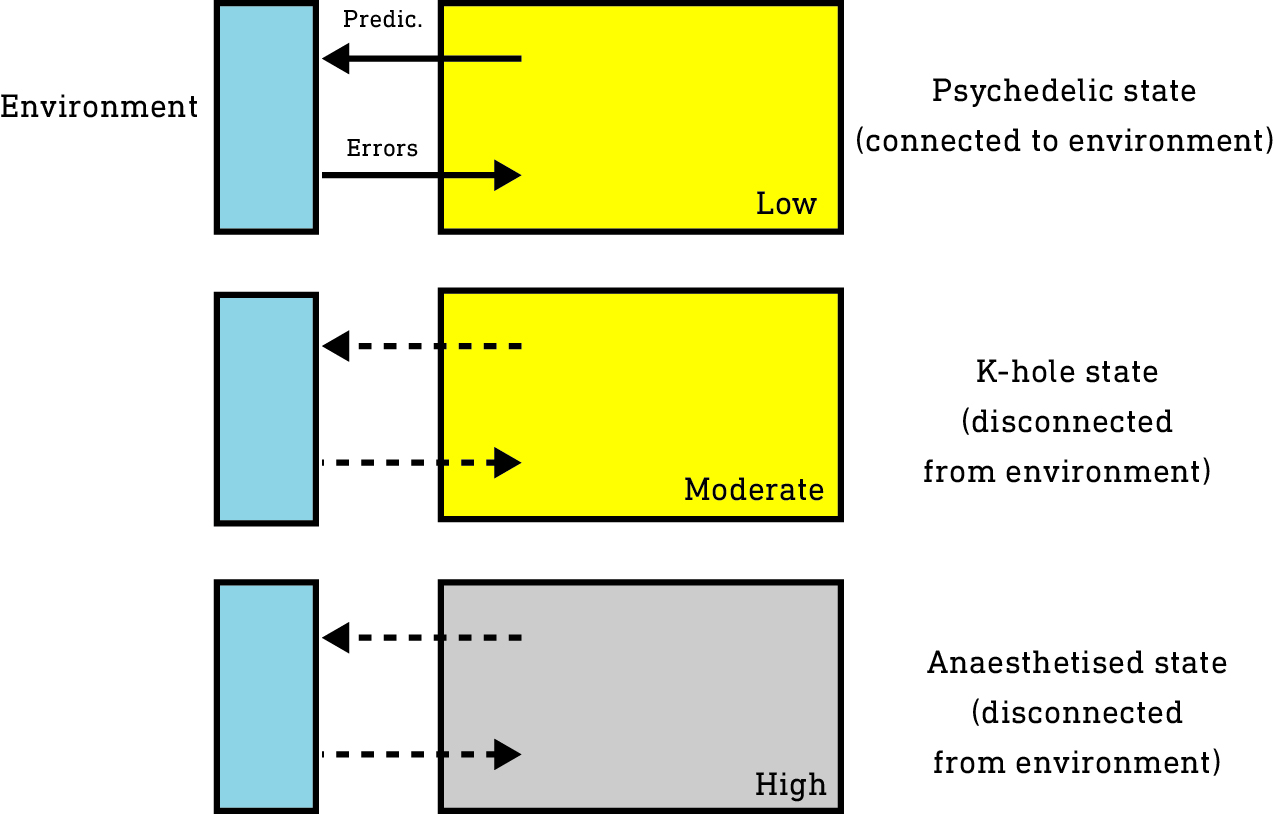
So, in summary, this model suggests a three-level effect plateau, rendering ketamine one of the more complex and, sometimes, unpredictable psychoactive molecules.
Low doses: cortical activation predominates resulting in a low-level psychedelic effect.
Moderate doses: cortical activation remains high, but prediction and error generating neurons are suppressed, disconnecting the world from the environment (the K-hole effect).
High doses: cortical activity is globally suppressed and the unique anaesthetised effect manifests.
For a much more detailed discussion and explanation of the neurological mechanisms behind the dissociative psychedelics, please refer to my latest book, Reality Switch Technologies: Psychedelics as Tools for the Discovery and Exploration of New Worlds, which covers all the major classes of psychedelics in unprecedented depth and detail. See my website for more details:
https://www.buildingalienworlds.com/books.html
